
| برتر |
| سلول |
| تابع |
| غشای سلولی سیستم غشایی داخلی غشای خارجی غشای دوگانه |
| هستی شناسی ژن |
|---|
| QuickGO |
بیوممبران یک لایه جداکننده است که محفظه سلولی را احاطه کرده یا به عنوان یک غشای سلولی ، داخل سلول را از بیرون جدا می کند. در داخل سلول، غشاهای زیستی با ساختارهای مختلف، اندامکها یا واکوئلها را از سیتوپلاسم جدا میکنند . یک بیوممبران، از طریق اجزای غشایی، نقش فعالی در انتقال انتخابی مولکول ها و انتقال اطلاعات بین دو محفظه ای که این بیوممبران بین آنها قرار دارد، دارد.
فهرست غشای زیستی بیوممبران
نفوذپذیری غشای زیستی بیوممبران
از آنجایی که غشای زیستی در درجه اول یک لایه جداکننده بین نواحی مختلف است، برای اکثر مولکول ها نفوذ ناپذیر است. مولکول های چربی دوست کوچکتر آزاد هستند تا در سراسر لایه های لیپیدی غشایی مانند دی اکسید کربن ، الکل ها و اوره منتشر شوند . به منظور ایجاد نفوذپذیری غشاء برای ذرات لیپوفوبیک مانند آب یا ذرات بزرگ مانند یون ها یا مولکول های قند ، پروتئین های حمل و نقل مختلفی در غشا تعبیه شده است که وظیفه انتقال مواد خاصی را بر عهده دارند. بنابراین از نفوذپذیری انتخابی صحبت می شود.
ساختار غشای زیستی بیوممبران
یک غشای زیستی همیشه از نظر توپولوژیکی بسته است و فضایی را در بر می گیرد. غشاهای غیرخودی در سلول های دست نخورده یافت نمی شوند. غشاهای زیستی نامتقارن هستند: آنها یک سمت پلاسماتیک (سمت P) رو به سیتوپلاسم و یک سمت خارج پلاسمیک (سمت E) دارند.
غشاهای زیستی از لیپیدها و پروتئین ها ساخته شده اند . زنجیره های کربوهیدرات را می توان به پروتئین ها متصل کرد. بخش لیپیدی ماده اصلی غشاء را به عنوان یک لایه دولایه لیپیدی تشکیل می دهد و مسئول خواص فیزیکوشیمیایی خاص آن است . به طور خاص، این لایه دوگانه به عنوان یک لایه جداکننده غیرفعال عمل می کند. استروئیدها مانند کلسترولوارد یک برهمکنش آبگریز با لیپیدها شده و در غلظتهای بالا در بیوممبران، غشای زیستی انعطافپذیر را تقویت میکند. علاوه بر این، پروتئین ها روی غشا و درون آن توزیع می شوند که عملکردهای فعال غشا را بر عهده می گیرند. پروتئین ها تنها عملکرد بسیار کوچکی از غشای زیستی دارند، زیرا در لایه های لیپیدی شنا می کنند.
بیوممبران ها را می توان با چگالی آنها مشخص کرد . معمولاً بین 1.12 و 1.22 گرم سانتیمتر -3 است . چگالی به نسبت وزنی پروتئین ها به لیپیدها بستگی دارد: بسته به عملکرد غشاء، مقادیر 0.25 ( غشای میلین ، محتوای پروتئین کم)، 1.3 (غشای پلاسمایی گلبول های قرمز )، 2.5 (غشای پلاسمایی E). coli )، 2.9 ( غشای میتوکندری داخلی ) تا مقدار 5 در غشای بنفش موجود در هالوباکتریوم (محتوای پروتئین بالا). [3]
در انواع خاصی از اندامک های سلولی ( هسته سلولی ، میتوکندری ، پلاستید )، غشاهای زیستی به صورت یک غشای دوتایی ظاهر می شوند .
دو لایه فسفولیپیدی غشای زیستی بیوممبران
دولایه لیپیدی عمدتاً از فسفولیپیدهای آمفیفیل تشکیل شده است که دارای یک گروه سر آبدوست و یک گروه دم آبگریز (عمدتاً زنجیرههای هیدروکربنی ) هستند. در آب، به عنوان یک نتیجه از اثر آبگریز ، یک دولایه تشکیل می شود که دم های آبگریز به سمت داخل و سرهای آب دوست نشان داده می شوند. به دلیل هسته آبگریز، چنین دولایه لیپیدی تقریباً در برابر آب و مولکول های محلول در آب نفوذ ناپذیر است، اما در عین حال بسیار انعطاف پذیر است و از نظر مکانیکی تخریب آن دشوار است. به همین دلیل، حتی سوراخ کردن با پیپت، سوراخی در غشا باقی نمیگذارد. برای این، می توان آن را با حلال های لیپیدی جایگزین کرد ولیپازها از بین می روند.
غشاها از سه نوع لیپید اصلی تشکیل شده اند: فسفوگلیسریدها، اسفنگولیپیدها و کلسترول.
فسفولیپیدها
- فسفولیپیدها با یک گروه فسفات مشخص می شوند ، آنها قسمت اصلی لیپیدهای غشا را تشکیل می دهند. آنها معمولاً یک ستون فقرات گلیسرول در سراسر غشاء دارند، به همین دلیل است که آنها فسفوگلیسرید نامیده می شوند. دو گروه از سه گروه هیدروکسی گلیسرول با اسیدهای چرب آبگریز استری می شوند و گروه سوم با یک گروه فسفات آبدوست. گروه فسفات می تواند جایگزین دیگری را حمل کند. اگر اینطور نباشد، که تقریباً در غشاها وجود ندارد، مولکول اسید فسفاتیدیک نامیده می شود . کولین ، که منجر به فسفاتیدیل کولین (PC) یا اتانول آمین می شود ، منجر به مکرر به عنوان یک جایگزین می شود.فسفاتیدیل اتانول آمین (PE)، سرین، باعث ایجاد فسفاتیدیل سرین (PS) یا اینوزیتول می شود ، و باعث ایجاد فسفاتیدیللینوزیتول (PI) می شود. این به کار میرود که تمام مولکولهای توصیفشده از یک گروه سر آبدوست، فسفات با جانشینها و یک دم آبگریز، یک اسید چرب بدون انشعاب با 16 تا 20 اتم تشکیل شدهاند. بسته به تعداد پیوندهای دوگانه در اسید چرب، بین اسیدهای چرب اشباع (بدون پیوند دوگانه )، تک غیراشباع (یک پیوند دوگانه) و اسیدهای چرب چند غیراشباع تمایز قائل میشوند.
اسفنگولیپیدها [ ویرایش | ویرایش منبع ]
- اسفنگولیپید ترکیبی از اسفنگوزین است که از طریق گروه آمینه آن به اسید چرب مرتبط است. گروه هیدروکسی را می توان با گروه های مختلف استری کرد ، بدون استری کردن سرامید می دهد ، استری با فسفوکولین اسفنگومیلین و با ساکاریدها گلیکوسفنگولیپید می دهد . اسفنگولیپیدها نیز آمفی پاتیک هستند و شبیه فسفولیپیدها هستند.
کلسترول غشای زیستی بیوممبران
- غشاهای حیوانات می توانند تا 50 درصد کلسترول ( درصد جرم ) داشته باشند ، در گیاهان کمتر و در باکتری ها اصلاً وجود ندارد. کلسترول کوچک است و خیلی آمفی پاتیک نیست، به همین دلیل است که فقط گروه هیدروکسی در سطح غشا و بقیه مولکول در غشاء قرار دارند. سیستم حلقه سفت و سخت کلسترول جریان لایه لیپیدی را مختل می کند و آن را سفت تر می کند. [4]

شماتیک دو لایه لیپیدی (مایع) یک غشای زیستی
دولایه لیپیدی یک غشای زیستی معمولاً مایع است. اچ. لیپیدها و پروتئین ها در سطح غشاء کاملاً متحرک هستند. با این حال، تبادل لیپیدها بین دو لایه یا حتی جدا شدن یک لیپید از غشاء بسیار نادر است. حرکت هدفمند از یک طرف غشاء به سمت دیگر ( فلیپ فلاپ ) معمولاً تنها با مشارکت فعال پروتئین های خاص (به اصطلاح فلیپازها و فلاپازها ) با استفاده از آدنوزین تری فسفات امکان پذیر است.(ATP) امکان پذیر است. فلیپازها لیپیدها را از بیرون غشای پلاسمایی به سمت سیتوزولی منتقل می کنند. فلوپازها ناقل ABC کلاسیک هستند و لیپیدهای غشایی را از سمت سیتوزولی غشای پلاسما به خارج منتقل می کنند. سایر ناقلین برای لیپیدهای غشایی اسکرامبلیسترها هستند که با این حال، لیپیدهای غشایی را در جهت گرادیان غلظتشان به روشی وابسته به ATP تا زمانی که تعادل برقرار نشود، مبادله نمی کنند.
اینکه دولایه لیپیدی چقدر سیال است در درجه اول به تعداد پیوندهای دوگانه در زنجیره های هیدروکربنی آبگریز لیپیدها بستگی دارد.برخی باکتری ها [5] نیز از شاخه های زنجیره ای استفاده می کنند. هرچه بیشتر باشد، غشا مایع بیشتری دارد. از طرف دیگر، درجه مایع توسط مولکول های تعبیه شده دیگر تعیین می شود. به عنوان مثال، کلسترول از یک طرف سیالیت را کاهش می دهد، اما از جامد شدن غشاء مانند ژل در دمای پایین جلوگیری می کند .
ویتامین E یک آنتی اکسیدان است (مانند ویتامین C )، از زنجیره های هیدروکربنی اشباع نشده فسفولیپیدهای غشای زیستی در برابر تخریب توسط رادیکال های آزاد ( پراکسیداسیون لیپیدی ) محافظت می کند.
پروتئین های غشای زیستی بیوممبران

مدل غشای سلولی بر اساس مدل موزاییک مایع
انواع مختلفی از پروتئینهای غشایی که در لایههای دوگانه لیپیدی تعبیه شدهاند، خواص متفاوت بیوممبرانها را از طریق برهمکنشهای پروتئین-لیپیدی تضمین میکنند. دو طرف یک غشای زیستی نیز به دلیل آرایش پروتئین های غشایی می توانند تفاوت زیادی داشته باشند. به عنوان مثال، در حالی که گیرنده های ارتباط سلول-سلول و برای تشخیص تغییرات محیطی به سمت بیرون اشاره می کنند، آنزیم های درگیر در واکنش ها به سمت داخل اشاره می کنند (یعنی در سیتوپلاسم قرار دارند).
بسیاری از پروتئین ها در حمل و نقل غشایی نقش دارند . اچ. در تبادل مواد و انتقال سیگنال از طریق گیرنده های خاص. تعداد زیادی از پروتئین های غشایی به خوبی مورد مطالعه قرار گرفته اند که انواع مختلف سلول و مراحل بلوغ آنها را مشخص می کنند و می توانند از فردی به فرد دیگر (مانند گروه های خون و بافت ) متفاوت باشند. این همچنین شامل مولکولهایی (عمدتاً گلیکوپروتئینها) میشود که به تمایز بین خود و خارجی طبق اصل قفل کلید کمک میکنند.
با توجه به مدل موزاییک مایع، پروتئین های غشایی به طور صلب در غشاء ثابت نیستند، اما قادر به تغییرات بسیار پویا در مکان درون غشا هستند. این پویاییها مبنایی برای راهاندازی زنجیرههای سیگنالینگ در سطح سلول، هم به صورت درون سلولی و هم بین سلولهای همکار تشکیل میدهند.
پروتئین های غشایی را می توان بر اساس لنگر انداختن آنها در دولایه لیپیدی طبقه بندی کرد:
پروتئین های انتگرال غشای زیستی بیوممبران
- توالی یابی ژن نشان می دهد که 30 درصد از تمام پروتئین های کدگذاری شده پروتئین های انتگرال هستند. پروتئین های انتگرال به صورت پروتئین های گذرنده عبور می کنندهر دو لایه دولایه لیپیدی، برخی تک، برخی دیگر با حلقه های متعدد. قسمت هایی از پروتئین از غشاء بیرون زده است. پروتئین های انتگرال، مانند فسفولیپیدها، آمفی پاتیک هستند. دامنههای درون غشاء آبگریز هستند، جایی که باقیمانده اسید آمینه با زنجیرههای لیپیدی تعامل دارد. با این حال، این نیروهای هدایت نشده به تنهایی برای تثبیت کافی نیستند. در بسیاری از پروتئینها، نواری از باقیماندههای عمدتاً پایه با سرگروههای باردار فسفولیپیدها در تعامل است. قسمت دیگر که از غشاء بیرون زده است، با آب اطراف و مواد محلول در آن برهم کنش دارد. پروتئین های انتگرال لزوماً به طور محکم در غشاء لنگر نیستند، اما می توانند آزادانه متحرک باشند.
پروتئین های محیطی
- پروتئین های محیطی را می توان در داخل و خارج غشاء قرار داد [6]. آنها به طور موقت به آنها یا به پروتئین های یکپارچه از طریق مخلوطی از برهمکنش های الکترواستاتیک و آبگریز و همچنین سایر پیوندهای غیر کووالانسی متصل می شوند. ضمیمه پویا است، بسته به شرایطی که میتوان آنها را بسته یا بیبند کرد. برای به دست آوردن آن لازم نیست غشاء از بین برود. یک محلول نمک بسیار غلیظ برای وارد کردن آنها به محلول کافی است، زیرا این امر برهمکنش های الکترواستاتیکی را ضعیف می کند. بهترین نمونه های مطالعه شده در سمت سیتوپلاسمی پروتئین هایی هستند که چیزی شبیه به اسکلت را به صورت فیبریل تشکیل می دهند، آنهایی که پوشش ها و آنزیم ها را تشکیل می دهند. پروتئین های محیطی بیرون عمدتاً به ماتریکس خارج سلولی تعلق دارند. پروتئین های یکپارچه و محیطی را می توان پس از ترجمه با اتصال به باقی مانده های اسید چرب، پرنیلاسیون اصلاح کرد.یا یک لنگر GPI .
پروتئین های لیپید لنگر
- پروتئین های لیپید لنگر جزو پروتئین های محیطی هستند و بنابراین از طریق غشاء نیز بیرون نمی زنند، اما به صورت کووالانسی به یک مولکول لیپیدی تعبیه شده در غشاء متصل می شوند. انواع مختلفی وجود دارد (به عنوان مثال پرنیلاسیون ( فارنسیلاسیون ، ژرانیل ژرانیلاسیون )، S-acylation یا myristoylation)، اما بسیاری از آنها لنگر GPI هستند . پروتئین هایی با لنگر GPI در قسمت بیرونی غشای پلاسمایی قرار دارند.
تابع
سیتوپلاسم داخل سلول توسط یک غشای زیستی از بیرون جدا می شود. اینها غشای سلولی ، غشای پلاسما ، پلاسمالما یا غشای سلولاریس نامیده می شوند . بیوممبران وظایف زیر را دارد:
- بخش بندی
- به دلایل انرژی، هر بیوممبران یک لایه پیوسته را نشان می دهد که با چندین غشاء، این امر به طور خودکار باعث ایجاد فضاهای جداگانه می شود که اصطلاحاً محفظه ها نامیده می شوند. بیشتر سلولها حاوی فضاهای واکنش و ذخیرهسازی ( قطعات )، مانند اندامکهای سلولی و واکوئلها با خواص شیمیایی بسیار متفاوت هستند. مواد بسیار متفاوتی در محفظه های مختلف وجود دارد. بنابراین، بسیار متفاوت، z. گاهی اوقات حتی فرآیندهای متضاد در همان زمان ممکن است که با یکدیگر تداخل نداشته باشند، مانند تجمع و تجزیه کربوهیدرات. علاوه بر این، تنظیم فردی امکان پذیر است.
- داربست برای فعالیت بیوشیمیایی
- برای واکنش های خاص، تراز دقیق مولکول ها نسبت به یکدیگر ضروری است، زیرا باید برهم کنش های خاصی وارد شود. این تراز دقیق در راه حل امکان پذیر نیست. اکنون بیوممبران ها داربستی را فراهم می کنند که روی آن مولکول ها می توانند به طور موثر با یکدیگر تعامل و واکنش نشان دهند. در غیر این صورت واکنش های مهم امکان پذیر نخواهد بود. برای مثال مجموعه چند آنزیمی زنجیره تنفسی و فتوسنتز در غشاء لنگر انداخته است.
- تراوایی انتخابی
- ذرات بدون مانع از غشاها عبور نمی کنند، اما می توانند انتخاب شوند و احتمالاً حفظ شوند.
- حمل و نقل مواد محلول
- مولکولها را میتوان از یک طرف غشاء به طرف دیگر، حتی در برابر یک گرادیان غلظت (یعنی به طور فعال) حمل کرد. به این ترتیب می توان مواد مغذی را در سلول غنی کرد. یون ها همچنین می توانند در سراسر غشاء منتقل شوند، این نقش مهمی برای اعصاب و عضلات دارد.
- پاسخ به سیگنال های خارجی
- غشای پلاسما برای پاسخ به محرک های خارجی (یعنی برای انتقال سیگنال) مهم است. گیرنده هایی در غشاء وجود دارد. اگر یک مولکول خاص در مجاورت آنها منتشر شود (یک لیگاند)، هر دو می توانند به دلیل ساختار مکمل خود ترکیب شوند و گیرنده سیگنالی را به سلول می فرستد. گیرندههای مختلف لیگاندهای مختلف را شناسایی میکنند و به سلول اجازه میدهند اطلاعاتی در مورد محیط خود دریافت کند. واکنشها به محیط، سازگاری متابولیسم با تغییر فعالیت آنزیم، آزادسازی مواد ذخیرهسازی یا حتی خودکشی است.
- تعامل بین سلولی
- غشای پلاسمایی لایه بیرونی سلول است. در موجودات چند سلولی، یک سلول از طریق غشای پلاسمایی با سلول های همسایه خود تعامل دارد. به عنوان مثال، سلول ها می توانند ب) به یکدیگر بچسبند یا سیگنال ها و مواد را مبادله کنند.
- تبدیل انرژی
- غشاها در تبدیل انرژی مانند فتوسنتز و تجزیه کربوهیدرات ها نقش دارند. در یوکاریوت ها، اولی در کلروپلاست ها و دومی در میتوکندری ها اتفاق می افتد .
- بزرگ شدن سطح
- برجستگی های کوچک غشای زیستی، به اصطلاح میکروویلی ، سطح سلول و در نتیجه ناحیه ای را که می توان روی آن کار کرد، بزرگ می کند، زیرا متابولیسم به ویژه بر روی غشای زیستی انجام می شود. [4]
سیالیت [ ویرایش | ویرایش منبع ]
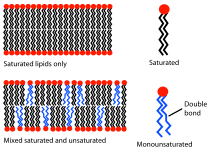
تاثیر اسیدهای چرب غیر اشباع بر ساختار غشا
سیالیت یک غشای زیستی به دما بستگی دارد . غشای فسفاتیدیل کولین و فسفاتیدیل اتانول آمین اشباع شده با بقایای اسیدهای چرب در دمای 37 درجه سانتیگراد کاملاً سیال خواهد بود. در این حالت، غشاء را می توان به عنوان یک کریستال مایع دو بعدی مشاهده کرد . محورهای بلند فسفولیپیدها به موازات یکدیگر قرار می گیرند، فسفولیپیدها خود می توانند بچرخند و آزادانه در صفحه حرکت کنند. تا دمای معین، دمای انتقال، حرکت فسفولیپیدها به شدت محدود می شود و خواص غشاء تغییر می کند، وضعیت اکنون بیشتر شبیه به حالت ژل منجمد است . دمای انتقال به ماهیت لیپیدها بستگی دارد. هر چه آنها کوتاهتر باشند و پیوندهای دوگانه بیشتر شوندآنها را شامل می شود، پایین تر است. کلسترول ساختار طبیعی غشاء را مختل می کند و تحرک لیپیدهای غشا را کاهش می دهد. دمای انتقال را دیگر نمی توان به طور واضح تعیین کرد. در سلول های حیوانی، کلسترول لیپیدی تضمین می کند که حالت مایع حتی با نوسانات دما حفظ می شود.
معنی [ ویرایش | ویرایش منبع ]
سیالیت یک غشای زیستی بین صلب و مایع قرار دارد و بنابراین ساختار خاصی را امکان پذیر می کند. پروتئین های غشایی می توانند در واحدهای عملکردی جمع شوند و بعداً دوباره جدا شوند. این برای مثال برای فتوسنتز مهم است. سیالیت همچنین نقش عمده ای در پیدایش غشاء ایفا می کند و برای بسیاری از فرآیندهای اساسی مانند تقسیم سلولی ، رشد سلولی ، ترشح و غیره مهم است. در حالی که دما اغلب در نوسان است، سیالیت غشاء باید ثابت بماند. به منظور دستیابی به این، لیپیدهای غشا را می توان اصلاح کرد: تبادل فسفولیپیدها امکان پذیر است. دساتورازهامی تواند پیوندهای دوگانه را از پیوندهای منفرد تشکیل دهد، ستون فقرات فسفات و دم لیپیدی فسفولیپیدها می توانند دوباره توزیع شوند و نسبت بیشتری از اسیدهای چرب غیراشباع نسبت به قبل تولید می شود. به این ترتیب، به ویژه موجودات خونسرد می توانند با محیط خود سازگار شوند.
قایق لیپیدی [ ویرایش | ویرایش منبع ]
مولکول های لیپید به طور یکنواخت در بیوممبران توزیع نمی شوند، اما میکرو دامنه هایی با ترکیب لیپیدی خاص وجود دارد. به ویژه کلسترول و اسفنگولیپیدها مستعد چنین ارتباطی هستند. برخی از پروتئین ها، مانند آنهایی که دارای لنگر GPI هستند، در چنین مناطقی تجمع می یابند، در حالی که برخی دیگر به ندرت در آنجا یافت می شوند. قایق های لیپیدی احتمالاً بسیار کوچک هستند و در یک روند دائمی انحلال و بازسازی هستند.
تاریخچه [ ویرایش | ویرایش منبع ]

نمودار دولایه لیپیدی با پروتئین های محیطی (مدل ساندویچی)

طرح آزمایشی آزمایش 1972 توسط فرای و ادیدین
- 1895 چارلز ارنست اورتون فرض می کند که غشاهای زیستی از لیپیدها تشکیل شده اند. او این را از مشاهدات به این نتیجه رسید که مواد چربی دوست (محلول در چربی)، به عنوان مثال داروهای بی حس کننده خاص، می توانند بسیار راحت تر از موادی که چربی گریز هستند وارد سلول ها شوند.
- 1917 ایروینگ لانگمویر پیشنهاد کرد که فسفولیپیدها روی سطح آب شناور هستند.
- در سال 1925 دانشمندان هلندی گوتر و گرندل مدل دولایه را توسعه دادند : [7] فسفولیپیدها با گروه های آبدوست به صورت یک لایه دولایه در غشاء قرار گرفته اند به گونه ای که گروه های آب دوست لیپیدها به سمت بیرون و گروه های آبگریز در داخل غشاء قرار می گیرند. دو لایه با این حال، این دو محقق با مدل خود محتوای پروتئین بزرگ غشاء زیستی را به طور کامل نادیده گرفتند.
- در سال 1935، JF Danielli و H. Davson مدل کلاسیک ساختار یک غشای زیستی را ارائه کردند: غشای زیستی از یک لایه لیپیدی دو مولکولی تشکیل شده است . دم های آبگریز لیپیدها رو به روی هم هستند، سرهای آبدوست با پروتئین پوشانده شده اند . به طور خلاصه: پروتئین – دولایه لیپیدی – پروتئین (ساختار ساندویچی). میکروگراف های الکترونی از غشاهای زیستی ساختار سه لایه ای را نشان می دهد: دو لایه بیرونی (هر کدام به ضخامت 2.5 نانومتر) و یک لایه میانی (ضخامت 3 نانومتر). این مدل غشایی یک غشای واحد نامیده می شود .
- در سال 1972، سیمور جاناتان سینگر و GL Nicolson مدل موزاییک سیال یک غشای زیستی را توسعه دادند : [8] مولکول های پروتئین کروی در یک فیلم لیپیدی دو مولکولی “شنا می کنند”. فیلم لیپیدی مانند یک مایع دو بعدی چسبناک عمل می کند ، به این معنی که مولکول های لیپیدی و پروتئین ها می توانند بدون مانع در صفحه غشاء پخش شوند . دو نوع ارتباط غشایی پروتئین ها وجود دارد. پروتئین های انتگرال که پروتئین های گذرنده نیز نامیده می شوند، از غشا عبور می کنند. پروتئینهای محیطی که پروتئینهای مرتبط نیز نامیده میشوند، در بالای دولایه لیپیدی قرار دارند.
- 1972: همزمان با سینگر و نیکلسون، فرای و ادیدین از آزمایشات روی دو سلول که در آن پروتئین های غشایی مشخص شده بودند، به این نتیجه رسیدند که غشاء نمی تواند ساکن باشد، اما در حرکت ثابت است. آنها سلول های مشخص شده و نواحی مشخص شده غشاء را که فقط به طور جداگانه وجود داشتند، با هم ترکیب کردند.
- در سال 1983 موریتسن و بلوم مدل تشک غشای سلولی را ارائه کردند. این بیان میکند که بخش آبگریز پروتئینهای غشایی که در غشاء تعبیه شدهاند، همیشه دقیقاً اندازه غشای سلولی ندارند و بنابراین لیپیدهایی با طول زنجیرههای مختلف به طور مناسب در اطراف پروتئینهای غشایی خاص ذخیره میشوند. [9]
- از زمانی که سینگر و نیکلسون مدل موزاییک مایع را در سال 1972 راهاندازی کردند، سرنخهای متعددی کشف شد که منجر به فرمولبندی مدل موزاییک با ساختار دینامیکی شد [10] . تحقیقات مختلف نشان داده است که پروتئین ها و مولکول های چربی مختلف به هیچ وجه به طور یکنواخت روی سطح غشا توزیع نمی شوند، همانطور که در یک مایع خالص انتظار می رود. در عوض، به نظر میرسد مناطقی با غلظت بالایی از پروتئینهای خاص (که جزایر گیرنده نامیده میشوند ) یا انواع خاصی از لیپیدها (که رافت لیپیدی نامیده میشوند) وجود دارد که دائماً در حال تنظیم مجدد، حل شدن و جمعآوری مجدد هستند.
سیستم غشای داخلی [ ویرایش | ویرایش منبع ]
سیستم غشای داخلی شامل بخشهای سلولی یا اندامکهای مختلف در سلولهای یوکاریوتی است که توسط غشاهای زیستی احاطه شدهاند. اینها عبارتند از: غشای هسته ای ، شبکه آندوپلاسمی ، دستگاه گلژی ، لیزوزوم ها ، وزیکول ها ، اندوزوم ها و غشای پلاسمایی . بیشتر پروتئینهایی که برای اندامکهای سیستم غشای داخلی تعیین میشوند، به صورت همزمان به شبکه آندوپلاسمی منتقل میشوند و از آنجا از طریق مسیر ترشحی در سیستم غشای داخلی منتقل میشوند .